y la evolución en las islas:
EL INTERESANTE CASO
DE HOMO FLORESIENSIS

Durante siglos, el ser humano se ha considerado una forma ajena a la naturaleza, libre para vivir y desarrollarse al margen de lo que las leyes naturales nos dicen. No obstante, poco a poco nos hemos visto en la necesidad de reconsiderar nuestras pretensiones al constatar, a través de los estudios científicos, que de ninguna forma existimos al margen de cómo funciona la vida en el planeta.
Desde hace muchos siglos, las islas han sido objeto de interés para el ser humano, pues no es extraño encontrar en su fauna y flora peculiaridades inesperadas: aves con impresionantes plumajes, flores gigantescas, insectos de descomunal tamaño, cangrejos que viven en tierra, mamíferos enanos, reptiles que nos recuerdan la era de los dinosaurios.
Estas peculiaridades responden a un básico y fundamental aspecto: la mayoría de las especies que habitan las islas se encuentran confinadas a ese espacio físico y geográfico. Muchas llegaron por azar: el viento arrastrando semillas o muy pequeños animales durante decenas y decenas de kilómetros; corrientes marinas en las que van semillas, animales como lagartijas o roedores flotando sobre troncos; aves o insectos que accidentalmente caen en una isla después de perder el rumbo o por algún accidente ocurrido; especies que ocupan una porción de tierra, un cabo o pequeña península,[1] la cual, por el cambio del nivel del mar, queda luego separada del continente y se convierte en isla. Ciertamente el azar juega un papel fundamental dentro del proceso de llegada de especies a una isla (figura 1), y lo demás depende de aspectos que a continuación veremos.
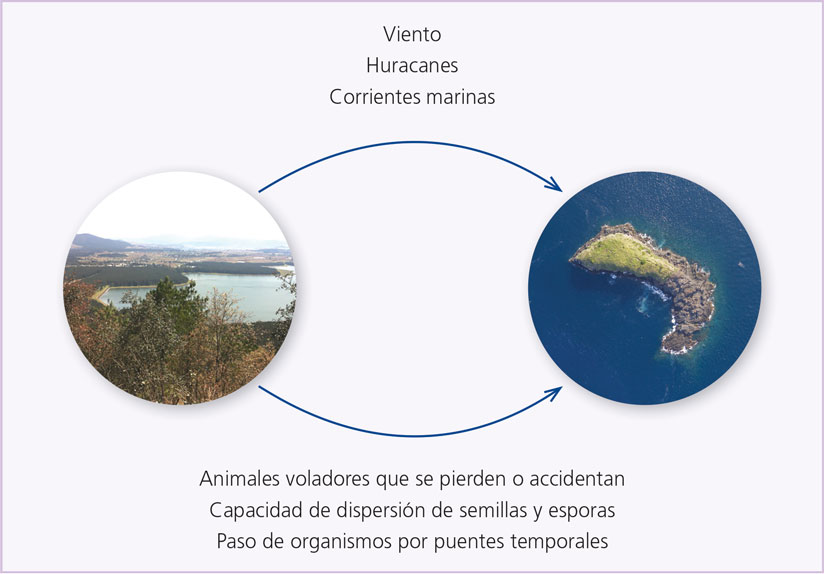
Figura 1. Formas de dispersión de organismos hacia islas (elaboración propia).
Salvo el último caso mencionado, la llegada de organismos a una isla por lo general implica que se enfrenten a una situación azarosa, pues las condiciones ambientales pueden ser enormemente diferentes a las de su hábitat original. Si se trata de individuos aislados, por ejemplo una araña llevada por el viento, quizá sobreviva; pero cuando muera, ahí terminará todo. No obstante, si se trata de semillas, por ejemplo, o de varios animales de la misma especie, entonces cabe la posibilidad de que, pese a la adversidad, sobrevivan y, si pueden reproducirse, den lugar a una nueva generación.
Aunque cada especie tiene características que la distinguen y además que la involucran con una forma de vida específica, los individuos de cada generación siempre tienen detalles que los diferencian de los demás: talla, color, flexibilidad alimentaria, conducta; si en un caso de aislamiento, alguna de estas características les ayuda a sobrevivir y reproducirse, eso les permitirá ser positivamente seleccionados y sus características tendrán más probabilidad de manifestarse de nuevo, en mayor cantidad e intensidad, generación tras generación. En realidad, estos casos de especies aisladas en islas son un muy claro ejemplo de selección natural y evolución (figura 2).
Veamos un caso muy significativo. El ave del paraíso (Paradisaea apoda) (figura 2) es una especie muy singular que habita en la isla de Nueva Guinea. El colorido de sus enormes plumas la distingue de inmediato, pero curiosamente este plumaje lo encontramos sólo en los machos y responde a un propósito: llamar la atención de las hembras para reproducirse.
Semejante condición podría afectarle a la hora de volar y alejarse de depredadores, y ciertamente sus alas son cortas, de modo que sólo se desplaza por el aire cuando realiza sus actos de cortejo o para pasar de un árbol a otro. Sin embargo, eso no es un problema, pues en esta isla no existen aves rapaces o mamíferos carnívoros arborícolas. Habita en la selva tropical y se alimenta de frutos e insectos.
Así como podemos encontrar numerosos casos de aves insulares que pierden sus habilidades de vuelo al encontrarse en ámbitos carentes de peligros, para el caso de los mamíferos es muy normal que la evolución lleve a la formación de especies de menor talla, pero con peculiaridades, por ejemplo: miembros proporcionalmente más cortos, cuerpos robustos pero de menor dimensión, por lo que los investigadores los definen no como chicos, sino más bien enanos. En el presente tenemos casos como el elefante pigmeo de Borneo (Elephas maximus borneensis), que habita dicha isla y cuya altura máxima es de 2.45 metros, poco en comparación con los casi tres metros de los ejemplares de la India.

Figura 2. Imágenes de Paradisaea apoda, una de las más interesantes y bellas aves del paraíso (familia Paradisaeidae). De una especie que ocupó las islas de Indonesia hace varios millones de años, derivaron las diferentes especies, muchas de ellas habitantes exclusivas de una sola isla. Ante la ausencia de depredadores y con el alimento seguro, la reproducción se convirtió en el principal factor de selección, lo que derivó en aves cuyos ejemplares masculinos son enormemente atractivos.
En 2004 se publicó un artículo en el que se describían restos fósiles de un hombre, de un individuo del género Homo, el cual había sido descubierto en la cueva de Liang Bua en la Isla de las Flores, en Indonesia (figura 3), y cuya antigüedad se calculó en unos 18 000 años antes del presente o incluso más (Brown, Sutikna, Morwood et al., 2004; Groves, 2007). En esa época el hemisferio norte vivía plenamente la época glaciar y las gigantescas masas de hielo retenían enormes cantidades de agua, situación que permitía que muchas islas estuvieran unidas a los continentes, lo cual facilitaba la dispersión de numerosas especies en esos territorios. Más tarde, cuando el clima cambió, los océanos aumentaron su volumen y así muchas penínsulas o cabos pasaron a ser islas, por lo que sus habitantes se quedaron atrapados.
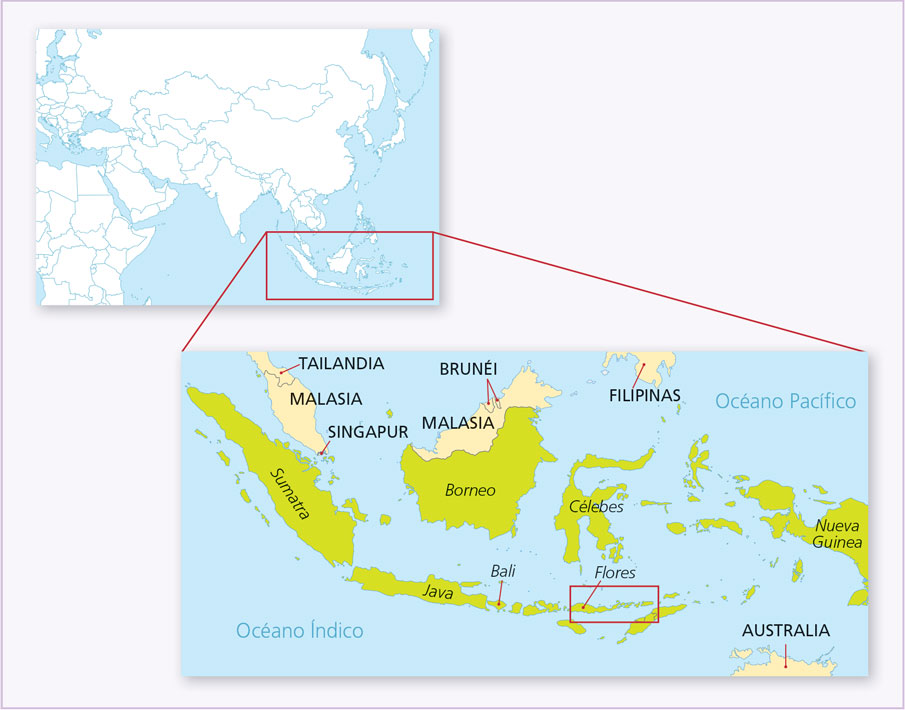
Figura 3. Mapa de Asia, región de Indonesia y, en el cuadro rojo, ubicación de la Isla de las Flores, lugar de hallazgo de Homo floresiensis (elaboración propia).
Los fósiles descubiertos correspondían a un adulto casi completo (denominado LB1), un simple premolar de otro individuo (LB2) (Groves, 2007) y numerosos instrumentos de piedra de minuciosa elaboración. El estudio de LB1 mostró que poseía frente huidiza, mentón muy chico y desvanecido y pequeños arcos superciliares (figura 4). Un detalle peculiar fue que sus piernas eran más bien cortas en comparación con los brazos, semejando un poco la morfología de los simios.[2] Es importante señalar también que en el mismo contexto se descubrieron restos fósiles de Stegodon, el ancestro directo de los elefantes modernos, pero en versión enana, y del dragón de Komodo (Varanus komodoensis).
Estos descubrimientos eran relevantes de por sí, pero sin duda los aspectos que más llamaron la atención fueron la pequeña estatura, ¡apenas un metro!, y –por si eso no era suficiente– la capacidad craneal para albergar un cerebro de ¡400 centímetros cúbicos!, es decir, casi una cuarta parte de lo que es el volumen del cerebro de Homo sapiens y casi igual que el de los actuales simios.
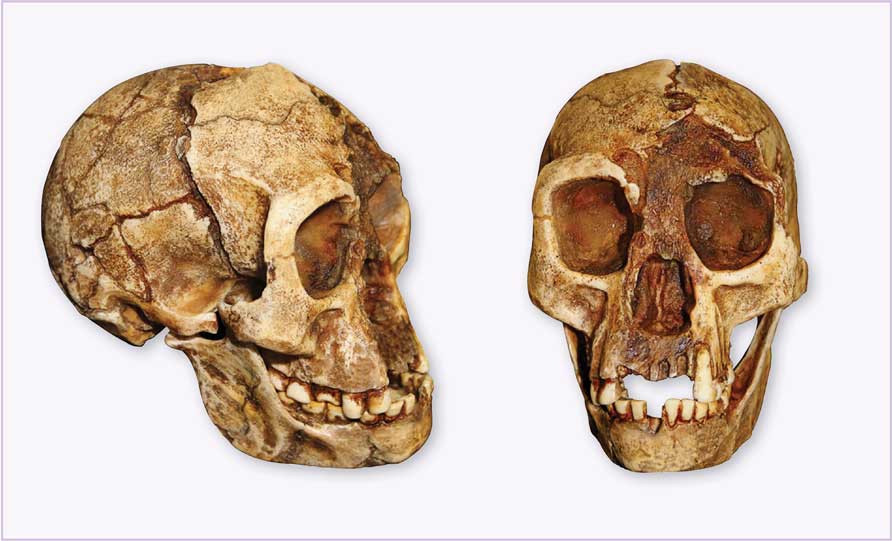
Figura 4. Imagen del cráneo de Homo floresiensis (individuo LB1) (Groves, 2007).
Los descubridores de estos fósiles fueron Thomas Sutikna, del Centro de Arqueología de Indonesia, en Jakarta, y Mike Morwood, de la Universidad de New England en Armidale (Nueva Gales del Sur), pero fue el paleoantropólogo Peter Brown quien los estudió, los describió, y ofreció las bases para proponerlos como nueva especie: Homo floresiensis. Brown propuso que se trataba de un caso de especiación por aislamiento insular y que era descendiente de Homo erectus, la especie de hombre más abundante y con mayor distribución en Asia hasta la llegada de Homo sapiens. Como dato curioso, en el año de su estudio (2004) estaba en su apogeo la serie de películas de El Señor de los Anillos, por lo que le dio al individuo más completo (LB1) el sobrenombre de “el hobbit”.
Sin duda este descubrimiento era de una magnitud equivalente al primer hallazgo de Australopithecus en las primeras décadas del siglo XX o del más antiguo integrante del género Homo: H. habilis, ya que nunca se había considerado la posibilidad de que pudieran darse modificaciones a la inversa en especies cercanas al hombre actual, es decir, tendencia a menor capacidad cerebral y a la formación de cuerpos que parecerían más del tipo de un chimpancé.
Como ocurre con frecuencia, pasada la sorpresa, gran parte del mundo paleoantropológico consideró necesario revisar toda la información disponible. Una de las propuestas más fuertes argumentaba que se trataba de un enano microcefálico del hombre actual o de Homo erectus, pero de ninguna manera de una especie diferente. Con base en ello se hicieron diversas investigaciones con el objetivo de comparar el cráneo de “el hobbit” (LB1) con casos microcefálicos de individuos de la región, sobre todo los que tenían además menor estatura, pero no se encontraron coincidencias en los detalles del cráneo ni se reconocieron patologías que son comunes en los casos de microcefalia.
Un aspecto que se notó en estos estudios es que, si bien el aislamiento insular podía dar lugar a especies de menor talla, difícilmente esto involucraba el tamaño del cerebro, pues incluso lo normal es que la masa cerebral sea proporcionalmente mayor respecto del cuerpo; por ejemplo, en el caso del tigre de Bengala (Panthera tigris tigris), habitante de la India, su peso puede rondar los 200 kilogramos, mientras que para el tigre de Sumatra (Panthera tigris sumatrae) apenas rebasa los 120. El tamaño del cerebro no varía entre uno y otro, de modo que el segundo posee un cerebro proporcionalmente más grande respecto del primero.
Cuadro 1. Capacidades craneales de simios y diversos homínidos
| Especie | Capacidad craneal (cc) |
|---|---|
| Chimpancé (Pan troglodytes) | 275-500 |
| Gorila (Gorilla gorilla) | 340-752 |
| Australopithecus spp. | 438-452 |
| Paranthropus spp. | 521-530 |
| Homo habilis | 612 |
| Homo erectus | 750-1300 |
| Homo neanderthalensis | 1100-1900 |
| Homo sapiens | 1200-1850 |
| Homo floresiensis | 400 |
Como puede verse, Homo floresiensis presenta un cerebro sumamente pequeño en comparación con las restantes especies de Homo (elaboración propia).
Siendo así el caso, ¿cómo entendemos los 400 centímetros cúbicos de capacidad craneal de Homo floresiensis? Este cuestionamiento derivó en una nueva alternativa: que esta especie era real, pero se había derivado de individuos patológicos de Homo erectus. Como quien juega un partido de tenis, pronto se tuvo una respuesta derivada del estudio del molde craneal de LB1, en el cual se concluyó que tenía un cerebro chico, sí, pero de estructura y morfología normales, a diferencia de la condición de microcefalia, la cual frecuentemente manifiesta cambios relevantes en su estructura. Un nuevo estudio publicado en 2008 involucró el análisis de un conjunto de medidas craneales de diversas especies, desde Australopithecus africanus hasta Homo sapiens (Lyras, Dermitzakis, Van der Geer et al., 2009) y la conclusión fue que H. erectus era la especie más afín.
Por último, investigaciones alrededor de la estructura mandibular manifestaron que sus características no son, de forma alguna, tipo Homo sapiens, sino más primitivas, lo cual dejó abierta la probabilidad de que su ancestro directo fuera Homo habilis, especie que habitó el oriente de África hace unos dos millones de años, aunque no existe ningún dato respecto de que alguna vez haya salido de este continente. Frente a dicha alternativa tenemos aquellas que lo ubican como descendiente de Homo erectus, quien desde hace millón y medio de años se distribuyó en casi toda Asia, incluyendo la isla de Java, donde sobrevivió hasta hace unos 100 000 años.
Para fortuna de la ciencia, las investigaciones en la cueva de Liang Bua continuaron y en 2005 hubo nuevos hallazgos, algunos correspondían a “el hobbit” (LB1) y otros eran piezas aisladas de un niño y seis adultos más. Todos muy acordes con lo observado antes (cuadro 2).
Cuadro 2. Colección de Homo floresiensis descubierta en la cueva
de Liang Bua, Isla de las Flores, Indonesia
| Individuos | Elementos descubiertos | Edad probable |
|---|---|---|
| LB1 | Esqueleto semicompleto | Adulto |
| LB2 | Molar aislado | Adulto |
| LB3 | Cúbito de pequeño tamaño | Adulto |
| LB4 | Radio y tibia | Adulto |
| LB5 | Vértebra y metacarpo | Adulto |
| LB6 | Mandíbula, escápula y varios metacarpos | Adulto |
| LB7 | Hueso del pulgar | Adulto |
| LB8 | Tibia | Adulto |
| LB9 | Fémur | Adulto |
Fuente: Groves 2007; elaboración propia
La temporalidad fue mucho más amplia de lo que se había determinado con los primeros hallazgos, pues varió entre los 95 000 y 12 000 años antes del presente. Esto permitió reconocer que la cueva de Liang Bua había sido ocupada de forma casi continua durante cerca de ¡60 000 años!
De igual manera, dentro de estas excavaciones aparecieron herramientas de piedra, pero desafortunadamente no estaban asociadas de modo directo con los individuos del clan hobbit, pues su antigüedad se ubicó sobre los 800 000 años. Este dato lleva a una significativa conclusión: que toda esta evolución desde Homo erectus hasta Homo floresiensis abarcó cerca de un millón de años, muy probablemente producto de la llegada del primero a la isla y de un aislamiento posterior que se prolongó por cientos de miles de años, tiempo suficiente para dar origen a una nueva especie.
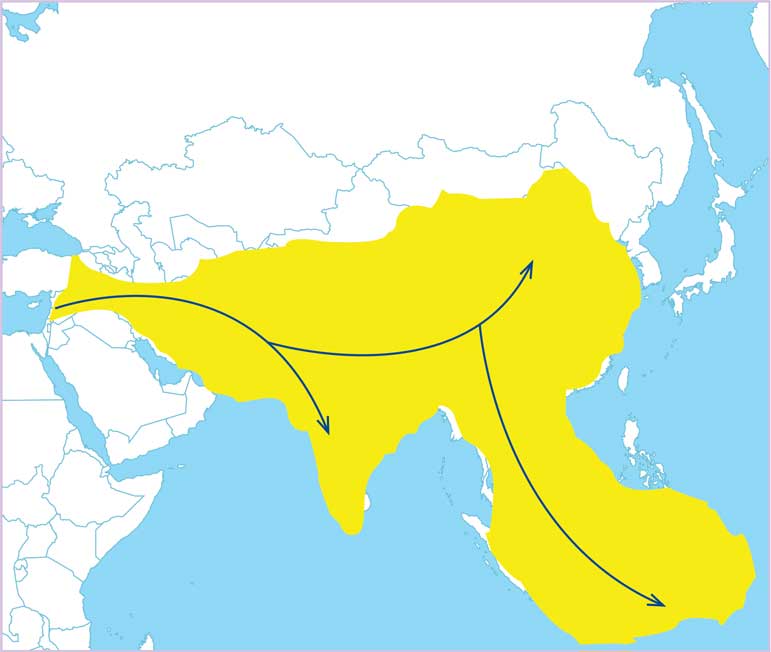
Figura 5. Distribución de Homo erectus y posibles rutas de dispersión (elaboración propia).
Un caso tan particular sin duda tiene muchos detalles que no pueden comprenderse aún, sobre todo si se quiere sostener la idea de que siendo este hombre un pariente no muy lejano de nosotros, necesariamente debía tener una serie de atributos, como la habilidad para hacer herramientas, usar el fuego y llevar una vida de cazador.
Pero lo cierto es que no en todos los casos existen elementos que lo confirmen. Por ejemplo, sabemos que elaboraba herramientas, pero no tenemos evidencia de que utilizara el fuego; por otro lado, frecuentemente se considera que la existencia de instrumentos de piedra significa habilidad cazadora, pero lo cierto es que la colección asociada refiere a actividades tales como golpear, perforar, cortar (Brumm, Aziz, Van den Bergh et al., 2006) y parece tener muy poca relación con puntas para sujetarlas a varas y así elaborar una lanza.
Un aspecto relevante es la presencia de brazos largos. Aunque no se habla mucho al respecto, en realidad se trata de una característica propia de los simios relacionada con el desplazamiento por los árboles (Tristán, 2004). Si bien es cierto que jamás se ha puesto en duda su condición bípeda, también lo es que por tratarse de ambientes de bosque tropical, la posibilidad de moverse entre las ramas sería de alto beneficio, sobre todo al buscar frutos, huevos o pequeños animales.
A partir de la información presentada podemos reconstruir la forma como vivió Homo floresiensis (figura 6). Hace unos 100 000 años, Homo erectus llegó a las islas del Asia Sudoriental, gracias a que algunas estaban unidas a tierra firme (figuras 7A y 7B). Al paso de los años, su presencia se afianzó e, incluso, gracias a la corta distancia pudo llegar a lugares que sí eran islas en el momento.
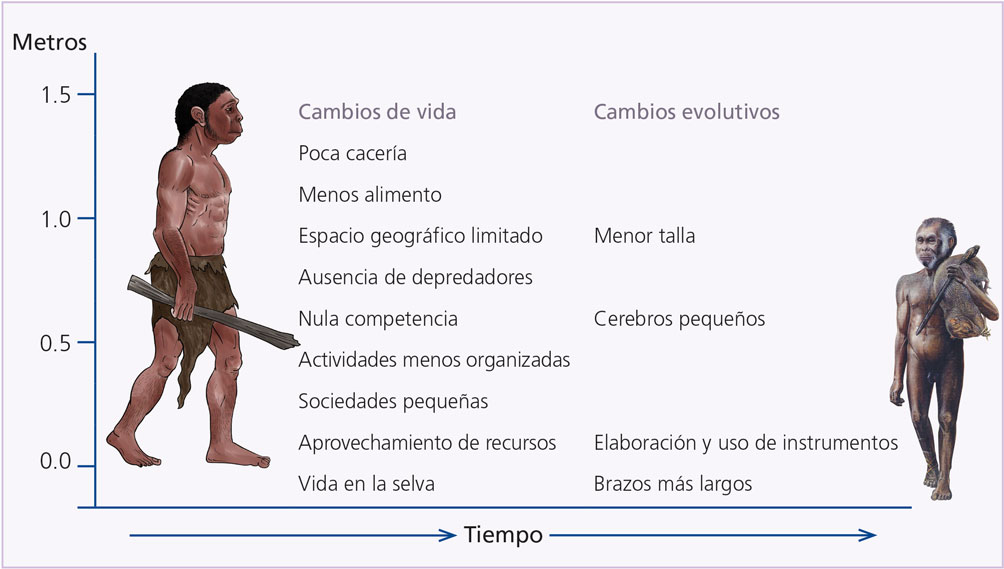
Figura 6. Evolución probable de Homo floresiensis a partir de Homo erectus. Su estatura, brazos comparativamente largos y pequeño cerebro quizá fueron el producto de la adaptación a la vida en una isla con vegetación de selva, sin depredadores ni competencia y con limitadas fuentes de alimento (elaboración propia).
Durante un tiempo, sus poblaciones iban y venían, aunque las que se encontraban en los extremos, por ejemplo en la Isla de las Flores, llegaban a condiciones de aislamiento y, por tanto, aparecieron detalles específicos, como la menor talla y los brazos más largos. Esta situación se mantuvo hasta hace unos 40 000 años, cuando tuvo lugar la llegada de Homo sapiens al continente asiático, pues con ello llevó a la extinción a su pariente H. erectus, e impulsó el aislamiento de las poblaciones que se encontraban en el extremo sudoriental (figura 7C). Para ese entonces, ya los hombres de la Isla de las Flores habían evolucionado hasta pasar a ser una especie nueva: Homo floresiensis, adaptada a la vida en bosques tropicales, apta para moverse en tierra y árboles, capaz de hacer herramientas para perforar, limpiar, golpear y matar mediante el impacto directo. Quizá porque fue un descubrimiento posterior o porque el ámbito tropical no lo hizo necesario, estos hobbits no manejaron el fuego.
Durante todo este periodo, las poblaciones de esta especie se vieron frente a un ambiente diverso, sí, pero limitado en espacio y carente de competencia o depredadores. Debido a ello, la selección natural actuó favoreciendo características como la menor talla. El aspecto del tamaño del cerebro no es algo muy claro, pero ciertamente el gasto energético de este órgano es enorme, por lo que también se favorecería la disminución de aquellas áreas que no eran fundamentales para sobrevivir, por ejemplo: habilidad para la elaboración de instrumentos sofisticados, lenguajes articulados o formación de grupos sociales complejos.
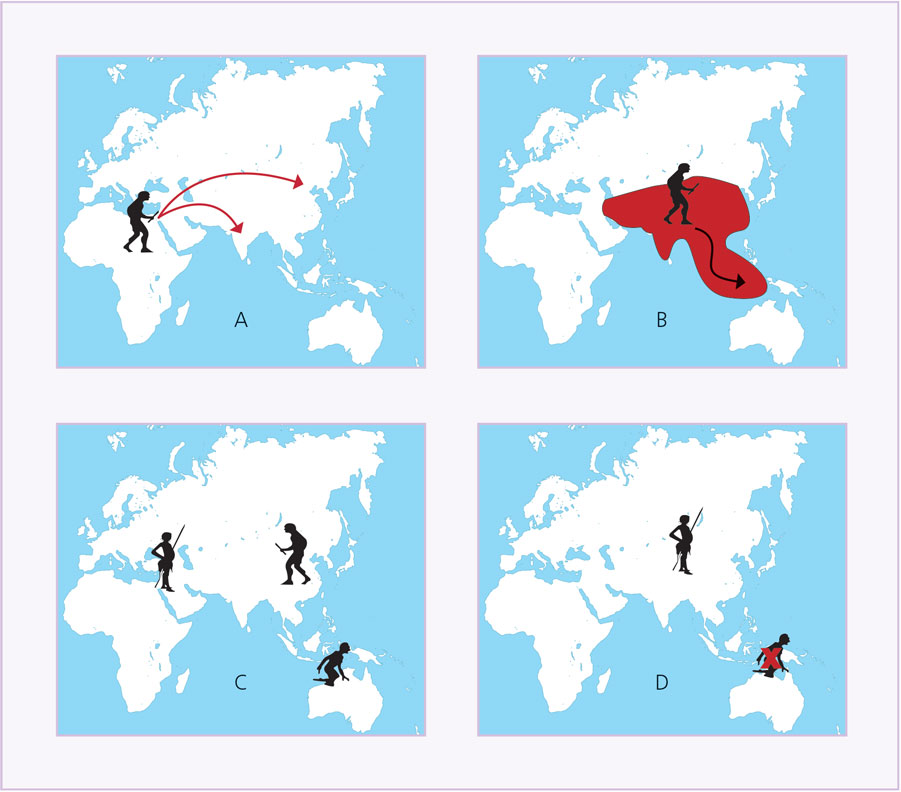
Figura 7. Sucesos claves en la historia de Homo floresiensis. Hace dos millones de años se originó Homo erectus (A) y de Africa pasó al continente asiático, ocupándolo y llegando hace unos 100 000 años al extremo de Asia Sudoriental (B). La distancia favoreció que las poblaciones más aisladas y alejadas evolucionaran, dando lugar a H. floresiencis, al tiempo que Homo sapiens avanza al interior de Asia, entrando en competencia con H. erectus (C) y llevándolo a la extinción. Hace 10 000 años H. sapiens se encuentra en toda Eurasia y África, y posiblemente provoca la desaparición de su pariente al avanzar rumbo a Australia (elaboración propia)
Aunque la lógica nos diga lo contrario, H. floresiensis vivió en esa isla durante más de 60 000 años. El motivo de su extinción no queda claro; algunos investigadores indican que fue producto de la actividad volcánica, otros que se debió a la llegada de nuestra especie a esa región, bien en época antigua o tan reciente como hace menos de un milenio (figura 7D). Comoquiera que sea, lamentablemente no tuvimos oportunidad de conocerlo y tenerlo como nuestra especie hermana.
El caso de esta peculiar especie de hombre nos permite ver un esquema diferente, interesante, de cómo puede funcionar la evolución en nuestro caso, llevando a algo diferente pero igualmente apto para sobrevivir y mantenerse por largo tiempo. Quizás haya quien niegue esto por la ineludible realidad de que se trata de una especie extinta, pero ante ello no hay respuesta mejor que la de su periodo de vida: casi 100 000 años. ¿Estamos seguros de que la civilización humana podrá romper esa marca? ♦
BROWN, P., T. Sutikna, M. J. Morwood, R. P. Soejono, Jatmiko, E. Wayhu Saptomo, y R. Awe Due (2004). A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. En: Nature, vol. 431, pp. 1055-1061.
BRUMM, A., F. Aziz, G. D. van den Bergh, M. J. Morwood, M. W. Moore et al. (2006). Early stone technology on Flores and its implications for Homo floresiensis. En: Nature, vol. 441, pp. 624-628.
GROVES, C. (2007). The Homo floresiensis Controversy. En: Journal of Biosciences, vol. 14, núm. 4, pp. 123-126.
LYRAS, G. A., M. Dermitzakis, A. van der Geer, S. B. van der Geer, y J. de Vos (2009). The origin of Homo floresiensis and its relation to evolutionary processes under isolation. En: Anthropological Science, vol. 117, núm. 1, pp. 33-43.
TRISTÁN, R. M. (2004). El homínido enano con cerebro de mono. En: El Mundo, vol. XV, núm., 5437, pp. 1-5 [en línea]: <www.almendron.com/tribuna/el-hominido-enano-con-cerebro-de-mono/>. Ir al sitio
Notas
* Doctor en Ciencias. Laboratorio de Paleozoología, Instituto de Investigaciones Antropológicas de la UNAM.- El nombre de cabo o punta, desde el punto de vista geográfico, corresponde a una porción de tierra de dimensiones pequeñas o moderadas que penetra en el mar.
- Grupo de primates al que pertenecen los chimpancés, gorilas, orangutanes y gibones.
- Imagen inicial: twitter.com/FossilHistory
- Foto 1: Dominio público en commons.wikimedia.org (CC BY-SA 4.0)
- Foto 2: carriekap.wordpress.com
- Foto 3: www.alamy.es
- Foto 4: Shutterstock
- Foto 5: movilpyme.com
CORREO del MAESTRO • núm. 291 • agosto 2020